Gezonde en zieke cellen 1 (2020/2021): werkgroep 5
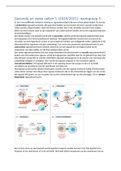
Er zijn 4 verschillende manieren waarop er signaaloverdracht bij een cel kan plaatsvinden. De eerste
is endocriene signaaloverdracht. Dit gaat doormiddel van hormonen die door speciale endocriene
cellen worden uitgescheiden. Deze hormonen worden dan in de circulatie vervoerd naar andere
delen van het lichaam waar ze op receptoren van cellen kunnen binden om zo een signaaloverdracht
te bewerkstelligen.
Een lokale manier van signaal overdracht is paracrien. Hierbij wordt het signaal uitgescheiden door
een bepaalde cel in het extracellulaire vloeistof. Het signaal blijft hier (wordt niet vervoerd in de
circulatie van het organisme) en kan zo op receptoren binden van omliggende cellen. (gebeurd zo bij
bijvoorbeeld het reguleren van een ontsteking). Een vorm van paracriene signaaloverdracht is ook
autrocriene signaaloverdracht (hierbij creeert de cel een signaal die vervolgens bindt op de
receptoren van deze zelfde cel: dit doen kankercellen soms).
Een ander signaaloverdracht dat over lange afstanden kan plaatsvinden is neurale signaaloverdracht.
Anders dan het endocriene signaal, wordt het signaal hier specifiek naar de cel gebracht ipv door het
hele lichaam verspreidt. Er worden elektrische signalen doorgegeven in de axonen van neuronen die
uiteindelijk eindigen in synapsen. Hier wordt het signaal omgezet in een chemisch signaal
(neurotransmitters). Dit signaal diffuseert in de opening tussen de synaps en de cel, en bindt
vervolgens op receptoren van het membraan van deze cel.
Het signaal op de kortste afstand heeft geen secretie van een signaal molecuul nodig. Hierbij maken
cellen direct contact met elkaar door signaal moleculen die in het celmembraan liggen van de cel die
het signaal wilt geven, en een receptor die op het celmembraan ligt van de ontvanger. Dit is contact-
dependent signaaloverdracht.
Een cel kan maar op een beperkt aantal signalen reageren omdat het niet voor alle signalen een
receptor in het membraan of in de cel heeft. Het heeft alleen receptoren op het membraan van de
, signalen die van belangrijk zijn voor de functie van die bepaalde cel. Afhankelijk van welk weefsel
zullen deze receptoren dus ook verschillen.
Zelfs als cellen dezelfde receptoren hebben voor hetzelfde signaal, reageren (response) ze niet
hetzelfde erop.
Hoe een cel weer reageert op een signaal hangt af van de set intracellulaire signaal moleculen die
elke cel heeft, en hoe deze moleculen de activiteit van effector eiwitten veranderen. Hierdoor
reageren verschillende cellen op een verschillende manier op het zelfde signaal.
Wanneer een hart pacemaker cel wordt blootgesteld aan acetylcholine, neemt de snelheid waarmee
hij afvuurt af. Maar wanneer een speeksel cel ook wordt blootgesteld aan acetylcholine, zal hij
onderdelen van speeksel gaan uitscheiden. Dit terwijl ze beide met dezelfde receptoren werken en
het zelfde signaal. Deze manier van anders reageren op hetzelfde signaal is handig omdat er maar 1
extracellulair signaal nodig is om veel verschillende effecten te hebben op verschillende weefsels;
denk bij voorbeeld aan de sympathicus en de parasympathicus.
Signaalroute van Acetylcholine.
Deze neurotransmitter bindt met een GPCR eiwit in het membraan van de cel. Dit geactiveerde
transmembraan eiwit, activeert nu de alfa-unit van een G-eiwit dat aan de cytosol-kant ligt van het
membraan (door middel van vorm veranderingen van het GPCR eiwit). Dit G-eiwit ( G q ) activeert op
zijn beurt weer het membraan gebonden enzym phospholipase C. Het signaal wordt hierna
doorgezet doordat phospholipase C een lipid molecuul dat onderdeel is van het celmembraan splijt;
dit is een inositol phospholipide (het is een phospholipide met een suiker inositol groep dat vast zit
aan zijn kop). Deze splijting resulteert in het ontstaan van twee second messenger moleculen: I P3
(inositol 1,4,5-triphosphate) en diacylglycerol (DAG) (dus de twee restproducten van het splitsen).
I P3 bindt in het cytosol met hierdoor open C a2+¿¿ kanalen in het membraan van het ER en SR. Zo
gaat er C a2+¿¿ vanuit het ER lumen naar het cytosol (deze stijging van C a2+¿¿ in het cytosol is ook
weer een signaal voor andere eiwitten; het activeert een PKC).
DAG blijft in het celmembraan na de splitsing. Hier werft het en activeert het eiwit kinase C (PKC) dat
zich eerst in het cytosol bevond (gaat nu naar celmembraan; wordt aangetrokken door DAG). Dit
enzym heeft ook C a2+¿¿ nodig om actief te worden. PKC kan geactiveerd een set van intracellulaire
eiwitten fosforyleren. (C a2+¿¿ komt ook vrij vanuit SR (sarcoplasmic reticulum) in spiercellen).
Er zijn 4 verschillende manieren waarop er signaaloverdracht bij een cel kan plaatsvinden. De eerste
is endocriene signaaloverdracht. Dit gaat doormiddel van hormonen die door speciale endocriene
cellen worden uitgescheiden. Deze hormonen worden dan in de circulatie vervoerd naar andere
delen van het lichaam waar ze op receptoren van cellen kunnen binden om zo een signaaloverdracht
te bewerkstelligen.
Een lokale manier van signaal overdracht is paracrien. Hierbij wordt het signaal uitgescheiden door
een bepaalde cel in het extracellulaire vloeistof. Het signaal blijft hier (wordt niet vervoerd in de
circulatie van het organisme) en kan zo op receptoren binden van omliggende cellen. (gebeurd zo bij
bijvoorbeeld het reguleren van een ontsteking). Een vorm van paracriene signaaloverdracht is ook
autrocriene signaaloverdracht (hierbij creeert de cel een signaal die vervolgens bindt op de
receptoren van deze zelfde cel: dit doen kankercellen soms).
Een ander signaaloverdracht dat over lange afstanden kan plaatsvinden is neurale signaaloverdracht.
Anders dan het endocriene signaal, wordt het signaal hier specifiek naar de cel gebracht ipv door het
hele lichaam verspreidt. Er worden elektrische signalen doorgegeven in de axonen van neuronen die
uiteindelijk eindigen in synapsen. Hier wordt het signaal omgezet in een chemisch signaal
(neurotransmitters). Dit signaal diffuseert in de opening tussen de synaps en de cel, en bindt
vervolgens op receptoren van het membraan van deze cel.
Het signaal op de kortste afstand heeft geen secretie van een signaal molecuul nodig. Hierbij maken
cellen direct contact met elkaar door signaal moleculen die in het celmembraan liggen van de cel die
het signaal wilt geven, en een receptor die op het celmembraan ligt van de ontvanger. Dit is contact-
dependent signaaloverdracht.
Een cel kan maar op een beperkt aantal signalen reageren omdat het niet voor alle signalen een
receptor in het membraan of in de cel heeft. Het heeft alleen receptoren op het membraan van de
, signalen die van belangrijk zijn voor de functie van die bepaalde cel. Afhankelijk van welk weefsel
zullen deze receptoren dus ook verschillen.
Zelfs als cellen dezelfde receptoren hebben voor hetzelfde signaal, reageren (response) ze niet
hetzelfde erop.
Hoe een cel weer reageert op een signaal hangt af van de set intracellulaire signaal moleculen die
elke cel heeft, en hoe deze moleculen de activiteit van effector eiwitten veranderen. Hierdoor
reageren verschillende cellen op een verschillende manier op het zelfde signaal.
Wanneer een hart pacemaker cel wordt blootgesteld aan acetylcholine, neemt de snelheid waarmee
hij afvuurt af. Maar wanneer een speeksel cel ook wordt blootgesteld aan acetylcholine, zal hij
onderdelen van speeksel gaan uitscheiden. Dit terwijl ze beide met dezelfde receptoren werken en
het zelfde signaal. Deze manier van anders reageren op hetzelfde signaal is handig omdat er maar 1
extracellulair signaal nodig is om veel verschillende effecten te hebben op verschillende weefsels;
denk bij voorbeeld aan de sympathicus en de parasympathicus.
Signaalroute van Acetylcholine.
Deze neurotransmitter bindt met een GPCR eiwit in het membraan van de cel. Dit geactiveerde
transmembraan eiwit, activeert nu de alfa-unit van een G-eiwit dat aan de cytosol-kant ligt van het
membraan (door middel van vorm veranderingen van het GPCR eiwit). Dit G-eiwit ( G q ) activeert op
zijn beurt weer het membraan gebonden enzym phospholipase C. Het signaal wordt hierna
doorgezet doordat phospholipase C een lipid molecuul dat onderdeel is van het celmembraan splijt;
dit is een inositol phospholipide (het is een phospholipide met een suiker inositol groep dat vast zit
aan zijn kop). Deze splijting resulteert in het ontstaan van twee second messenger moleculen: I P3
(inositol 1,4,5-triphosphate) en diacylglycerol (DAG) (dus de twee restproducten van het splitsen).
I P3 bindt in het cytosol met hierdoor open C a2+¿¿ kanalen in het membraan van het ER en SR. Zo
gaat er C a2+¿¿ vanuit het ER lumen naar het cytosol (deze stijging van C a2+¿¿ in het cytosol is ook
weer een signaal voor andere eiwitten; het activeert een PKC).
DAG blijft in het celmembraan na de splitsing. Hier werft het en activeert het eiwit kinase C (PKC) dat
zich eerst in het cytosol bevond (gaat nu naar celmembraan; wordt aangetrokken door DAG). Dit
enzym heeft ook C a2+¿¿ nodig om actief te worden. PKC kan geactiveerd een set van intracellulaire
eiwitten fosforyleren. (C a2+¿¿ komt ook vrij vanuit SR (sarcoplasmic reticulum) in spiercellen).