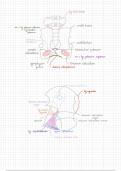
In de kern van elke cel ligt erfelijk materiaal (DNA) opgeslagen in de vorm van
chromosomen. Alle erfelijke informatie komt twee keer voor: één chromosoom van de
moeder en één chromosoom van de vader.
Er zijn 23 chromosomenparen:
- 22 paar autosomale chromosomen
- één paar geslachtschromosomen
- XX voor een meisje
- XY voor een jongen
(geslacht wordt bepaald door welk geslachtschromosoom de vader afgeeft)
DNA bestaat uit drie miljard basenparen en 20678 genen, dit is 1,2% van het genoom.
Veranderingen in DNA kunnen leiden tot veranderingen in eiwitten, die de oorzaak kunnen
zijn van verschillende aandoeningen. Want eiwitten hebben functies, zoals het kunnen laten
plaatsvinden/regelen van processen in het lichaam.
Eiwitsynthese
1. transcriptie: DNA → pre RNA
2. splicing: pre RNA → mRNA (introns worden verwijderd, exons blijven over)
3. translatie: mRNA → eiwitten (mRNA wordt afgelezen door ribosomen in het
cytoplasma, waardoor een polypeptideketen wordt gevormd)
Er zijn drie manieren om een stukje DNA af te lezen (reading frames / leesramen), maar
alleen het leesraam met het startcodon op de juiste plek wordt gebruikt.
DNA-MUTATIES
Een mutatie is pathogeen als:
- deze de functie van het eiwit waarvoor het hoort te coderen vermindert
- deze niet in gezonde personenen te vinden is
- patiënten met mutatie dezelfde symptomen of klinische kenmerken hebben
- familieleden met de mutatie de ziekte ook hebben
puntmutatie = één nucleotide verandert in een andere (A verandert bijv in een T)
- silent: het gemuteerde triplet/codon codeert alsnog voor hetzelfde aminozuur, dus
geen verandering in het eiwit → niet pathogeen
- missense: het triplet/codon is zodanig veranderd dat het codeert voor een ander
aminozuur en dus voor een verandering in het eiwit → niet altijd pathogeen
- nonsense: de triplet/codon is zodanig veranderd dat het een stopcodon is geworden
→ vrijwel altijd pathogeen
- SNP (single nucleotide polymorphism): een nucleotide die variabel is tussen
individuen: op een plaats in het genoom kan men bij verschillende mensen een
ander nucleotide aantreffen, er wordt bijvoorbeeld van een SNP gesproken als de
DNA-volgorde op een bepaalde plaats op een chromosoom zowel AAGCCTA als
AAGCTTA kan zijn.
SNPs hebben vaak geen effect op het fenotype of de functie van eiwitten en zijn
meestal onschuldig, doordat een heel groot deel in de introns ligt. Zelfs als een SNP
in een exon ligt hoeft dit niet te leiden tot een verandering in het eiwit:
, - silent → niet pathogeen
- missense mutaties in exons
- nonsense mutaties in exons
SNPs buiten de exons kunnen alsnog leiden tot een veranderingen in eiwitten:
- SNP treedt op in een stuk DNA dat essentieel is voor de transcriptieregulatie,
zoals sequenties die functioneren als promotor (aan het begin van
genen/exons die ervoor zorgt dat de transcriptie daar begint, markeert de
start) / enhancer (rekruteert activatie eiwitten, die ervoor zorgen dat RNA
polymerase efficiënt verloopt) → bijna altijd pathogeen
- SNP treedt op in een nucleotide die cruciaal is in een splice site, door de
mutatie kan een exon worden overgeslagen of kan frameshift optreden,
waardoor te vroeg een stopcodon ontstaat → bijna altijd pathogeen
insertie: toevoeging van één of meer nucleotiden/triplets
deletie: verwijdering van één of meer nucleotiden/triplets
Als er sprake is van insertie/deletie van hele tripletten, wordt een heel aminozuur verwijderd
of toegevoegd. Dan is de insertie/deletie in frame.
Als er geen sprake is van insertie/deletie van hele tripletten, maar van enkele nucleotiden,
dan is er sprake van frameshift (leesraamverschuiving): na de mutatie worden compleet
andere aminozuren afgelezen en ontstaan dus ook compleet andere eiwitten → vrijwel altijd
pathogeen.
Bij splice-site mutaties is er een mutatie opgetreden in nucleotiden die zich aan de
uiteinden van intronen bevinden en exonherkenning stimuleren. Zo’n mutatie kan ervoor
zorgen dat een exon niet wordt opgenomen in het mRNA of dat een (deel van) een intron
juist wel in het mRNA wordt opgenomen → bijna altijd pathogeen
- splice acceptor variant zit vóór een exon en leidt tot het overslaan van het
volgende exon
- splice donor variant zit altijd na een exon en leidt tot het overslaan van het vorige
exon
Planten hebben een stuk meer genen dan dieren en mensen, doordat ze zelf veel stoffen
moeten aanmaken en door alternatieve splicing: hierdoor kunnen we (mensen en dieren)
van dezelfde exonen verschillende mRNA’s maken die coderen voor verschillende eiwitten
en hebben we dus minder genen nodig.
Human genome reference sequence is een perfecte DNA-sequentie zonder pathogene
varianten, die we gebruiken om in het genoom van zieke mensen de pathogene varianten te
herkennen = gouden standaard (1 op 1000 nucleotiden van een gezond met is anders dan
de gouden standaard).
De genomische variant wordt als pathogenetisch beschouwd als deze aan de volgende
voorwaarden voldoet:
1. de variant vermindert de functie van het eiwit waarvoor het hoort te coderen
2. de variant komt niet voor bij gezonde individuen
3. patiënten met dezelfde pathogenetische variant hebben dezelfde symptomen of
klinische kenmerken
chromosomen. Alle erfelijke informatie komt twee keer voor: één chromosoom van de
moeder en één chromosoom van de vader.
Er zijn 23 chromosomenparen:
- 22 paar autosomale chromosomen
- één paar geslachtschromosomen
- XX voor een meisje
- XY voor een jongen
(geslacht wordt bepaald door welk geslachtschromosoom de vader afgeeft)
DNA bestaat uit drie miljard basenparen en 20678 genen, dit is 1,2% van het genoom.
Veranderingen in DNA kunnen leiden tot veranderingen in eiwitten, die de oorzaak kunnen
zijn van verschillende aandoeningen. Want eiwitten hebben functies, zoals het kunnen laten
plaatsvinden/regelen van processen in het lichaam.
Eiwitsynthese
1. transcriptie: DNA → pre RNA
2. splicing: pre RNA → mRNA (introns worden verwijderd, exons blijven over)
3. translatie: mRNA → eiwitten (mRNA wordt afgelezen door ribosomen in het
cytoplasma, waardoor een polypeptideketen wordt gevormd)
Er zijn drie manieren om een stukje DNA af te lezen (reading frames / leesramen), maar
alleen het leesraam met het startcodon op de juiste plek wordt gebruikt.
DNA-MUTATIES
Een mutatie is pathogeen als:
- deze de functie van het eiwit waarvoor het hoort te coderen vermindert
- deze niet in gezonde personenen te vinden is
- patiënten met mutatie dezelfde symptomen of klinische kenmerken hebben
- familieleden met de mutatie de ziekte ook hebben
puntmutatie = één nucleotide verandert in een andere (A verandert bijv in een T)
- silent: het gemuteerde triplet/codon codeert alsnog voor hetzelfde aminozuur, dus
geen verandering in het eiwit → niet pathogeen
- missense: het triplet/codon is zodanig veranderd dat het codeert voor een ander
aminozuur en dus voor een verandering in het eiwit → niet altijd pathogeen
- nonsense: de triplet/codon is zodanig veranderd dat het een stopcodon is geworden
→ vrijwel altijd pathogeen
- SNP (single nucleotide polymorphism): een nucleotide die variabel is tussen
individuen: op een plaats in het genoom kan men bij verschillende mensen een
ander nucleotide aantreffen, er wordt bijvoorbeeld van een SNP gesproken als de
DNA-volgorde op een bepaalde plaats op een chromosoom zowel AAGCCTA als
AAGCTTA kan zijn.
SNPs hebben vaak geen effect op het fenotype of de functie van eiwitten en zijn
meestal onschuldig, doordat een heel groot deel in de introns ligt. Zelfs als een SNP
in een exon ligt hoeft dit niet te leiden tot een verandering in het eiwit:
, - silent → niet pathogeen
- missense mutaties in exons
- nonsense mutaties in exons
SNPs buiten de exons kunnen alsnog leiden tot een veranderingen in eiwitten:
- SNP treedt op in een stuk DNA dat essentieel is voor de transcriptieregulatie,
zoals sequenties die functioneren als promotor (aan het begin van
genen/exons die ervoor zorgt dat de transcriptie daar begint, markeert de
start) / enhancer (rekruteert activatie eiwitten, die ervoor zorgen dat RNA
polymerase efficiënt verloopt) → bijna altijd pathogeen
- SNP treedt op in een nucleotide die cruciaal is in een splice site, door de
mutatie kan een exon worden overgeslagen of kan frameshift optreden,
waardoor te vroeg een stopcodon ontstaat → bijna altijd pathogeen
insertie: toevoeging van één of meer nucleotiden/triplets
deletie: verwijdering van één of meer nucleotiden/triplets
Als er sprake is van insertie/deletie van hele tripletten, wordt een heel aminozuur verwijderd
of toegevoegd. Dan is de insertie/deletie in frame.
Als er geen sprake is van insertie/deletie van hele tripletten, maar van enkele nucleotiden,
dan is er sprake van frameshift (leesraamverschuiving): na de mutatie worden compleet
andere aminozuren afgelezen en ontstaan dus ook compleet andere eiwitten → vrijwel altijd
pathogeen.
Bij splice-site mutaties is er een mutatie opgetreden in nucleotiden die zich aan de
uiteinden van intronen bevinden en exonherkenning stimuleren. Zo’n mutatie kan ervoor
zorgen dat een exon niet wordt opgenomen in het mRNA of dat een (deel van) een intron
juist wel in het mRNA wordt opgenomen → bijna altijd pathogeen
- splice acceptor variant zit vóór een exon en leidt tot het overslaan van het
volgende exon
- splice donor variant zit altijd na een exon en leidt tot het overslaan van het vorige
exon
Planten hebben een stuk meer genen dan dieren en mensen, doordat ze zelf veel stoffen
moeten aanmaken en door alternatieve splicing: hierdoor kunnen we (mensen en dieren)
van dezelfde exonen verschillende mRNA’s maken die coderen voor verschillende eiwitten
en hebben we dus minder genen nodig.
Human genome reference sequence is een perfecte DNA-sequentie zonder pathogene
varianten, die we gebruiken om in het genoom van zieke mensen de pathogene varianten te
herkennen = gouden standaard (1 op 1000 nucleotiden van een gezond met is anders dan
de gouden standaard).
De genomische variant wordt als pathogenetisch beschouwd als deze aan de volgende
voorwaarden voldoet:
1. de variant vermindert de functie van het eiwit waarvoor het hoort te coderen
2. de variant komt niet voor bij gezonde individuen
3. patiënten met dezelfde pathogenetische variant hebben dezelfde symptomen of
klinische kenmerken