Cellulaire fysiologie : hoofdstuk 2 : membraan
structuur
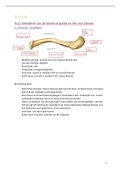
1. Oppervlak van de cel wordt gedefinieerd door een membraan
Opbouw:
• Fosfolipidendubbellaag door amfipatisch karakter:
o Hoofd = polair/ hydrofield (wijst naar het water)
o VZ staart = apolair/hydrofoob (wijst weg van het water)
• Ondoorlaatbaar voor grote moleculen (proteïnen en NZ)
• Selectief permeabel voor kleine moleculen (ionen en metabolieten)
• Actief transport doorheen het membraan = stoffen worden tegen hun concentratiegradiënt
in via een membraan-eiwit (ATP) opgenomen in de cel of afgestaan aan het milieu. (stoffen
worden dus opgenomen vanuit een gebied met een lage conc -> hoge conc)
2. Celmembraan is opgebouwd uit fosfolipiden
Fosfolipidenmembraan vormt in laag geconcentreerd waterig milieu een monolaag
En in een hoog geconcentreerd waterig milieu micellen
Bij een nog hogere concentratie aan water wordt een fosfolipidedubbelllaag gevormd = bilayer
Leaflets = fosfolipidemoleculen liggen in 2 parallelle rijen staart à staart tegen elkaar
Bilayer:
-> spontane vesikelvorming
-> spontaan helen bij ruptuur (inscheuren van weefsel)
-> self-sealing: kan geen vrije rand hebben
-> spontane opvouwing/regeneratie
1
,Meeste membraanlipiden = fosfolipiden (bv. fosfatiylcholine)
• Hoofd = hydrofiel = glycerol – fosfaatgroep – choline
• Staart = hydrofoob = 2 acylstaarten
• Verzadigd/ saturated= recht
• Onverzadigd/ unsaturated = knik t.h.v. dubbele binding
• Verzadigheid/onverzadigdheid beïnvloednen de VL, dikte en structuur van het membraan
• Fosfolipiden van dierlijke cellen bevatten meestal 1 verzadigde en 1 onverzadigde VZ staart
Andere membraanlipiden
- cholesterol
- glycolipiden: fosfatidylserine, fosfatidylinositol
- sfingolipiden (afkomstig van sfingosine) : glycosfingolipiden, sfingomyeline, gangliosides
3. Bilayer vormt een 2D vloeibare flase
Beweging van lipiden in de fosfolipidendubbellaag:
• Laterale diffusie = snel
• Rotatie/flexie van lipide = snel
• Flip-flop beweging van de ene -> andere leaflet = traag en gebeurd nauwelijks -> hoge
energie barrière verreist en floppase enzymen
Flipase = fosfolipiden van EC -> IC brengen
Flopase = fosfolipiden van IC -> EC brengen
Scramblase = fosfolipiden van EC -> IC en IC -> EC brengen (kan beide)
4. Vloeibaarheid van het membraan wordt bepaald door lipiden compositie
1. Zuiver fosfolipide membraan: transitietemperatuur hangt af van samenstelling
(packing) membraan:
• Lengte fosfolipiden staarten: 14-24C (meeste 18-20), hoe korter de staart -> hoe
vloeibaarder het zuiver fosfolipide membraan
• Aantal onverzadigdheden in lipide staarten:
o Verzadigd: recht -> dichte/sterke packing
o Onverzadigd: knik -> minder/zwakke packing -> vloeibaar bij lagere temperatuur
o Gevolg (nadeel) van zwakke packing is dat het membraan scheuren/gaten vertoont
en dus meer permeabel is
• Korte verzadigde ketens -> lage Tm (bv. olie)
• Lange onverzadigde ketens -> hoge Tm (bv. boter)
Tm = transitietemperatuur = temperatuur waarbij de
bilayer overgaat van de vaste ‘gel’ fase naar de vloeibare
‘sol’ (en omgekeerd)
2
, 2. Biologisch membraan:
• Zelfde algemene principes maar meer soorten lipiden
• Cholesterol intercaleerd met onverzadigde lipiden en heeft 2 effecten:
o Rigide ring stabiliseert C-atomen nabij ‘head’ -> rigider, minder permeabel
o Cholesterol verhindert staart-staart interactie (voorkomt kristallisatie van fosfolipide
staarten) -> membraan blijft vloeibaar
o Invloed op dikte van het membraan -> wordt dikker
hoge concentratie aan cholesterol -> vloeibaarheid membraan stijgt
in tegenstelling tot fosfolipiden gebeuren cholesterol
flipflops makkelijk over het membraan omdat
cholesterol een polair hoofdje heeft
-> gelijke concentratie aan cholesterol aan de binnen
en buitenkant van het membraan
Stijfheid/ vloeibaarheid van de bilayer bepaald de
permeabiliteit van het membraan
5. Fosfolipidedubbellaag is ondoorlaatbaar voor geladen moleculen
Zuivere fosfolipidedubbellaag is ondoorlaatbaar voor
-> ionen: Na+, K+, Cl-, Ca+
-> grote wateroplosbare moleculen: proteïnen, NZ, suikers en nucleotiden
Permeabel voor:
-> O2, CO2 en NH3
6. Asymmetrie in biologische membranen
Inner leaflet (intracellulair): Outer leaflet (extracellulair):
- fosfatidylinositol - glycolipiden
- fosfatidylethanolamine - sfingomyeline
- fosfatidylserine - fosfatidylcholine
Fosfatidylinositol -> betrokken bij second messenger signaal: IP3-DAG/PKC signaal cascade -> IC
Glycolipiden enkel aan de extracellulaire zijde: door synthese in ER/ golgi lumen
Fosfatidylchoine wordt in de binnenzijde aangemaakt en via flopase naar buitenzijde
3
,Aanmaak van fosfolipiden in SER of GA in lumen of in de cytoplasmatische zijde
Asymmetrie in fosfolipide compositie tussen inner en outer leaflet van het membraan:
• Ontstaan tijdens biosynthese (ER/golgi: flippase/floppase)
• Beïnvloed de buiging en vloeibaarheid van het membraan (EC is stijver dan IC)
• Cytosol leaflet (IC) is negatiever dan EC, reguleert incorporatie van membraan proteïnen
o Fosfatidylserine heeft een negatieve lading, de andere FL zijn neutraal
Specifieke compositie is soms nodig voor de functie van membraan proteïnen
7. Lipid rafts
Lipid raft = locale concentratie van specifieke lipiden, sfingomyeline, cholesterol en proteïnen
• Meestal dikker dan de rest van het membraan door aanrijking van verzadigde fosfolipiden
• Clusters van proteïnen/lipiden (rafts) spelen een rol in signaaltransductie (concentratie
second messengers zoals PIP3)
• Bevatten glycolipids (geglycosyleerde fosfolypiden
8. Correct incorporeren van transmembraanproteïnen
8.1. Transmembraanproteïnen
Soorten:
- perifere proteïnen zijn niet covalent gebonden met integrale proteïnen
- meeste integrale membraaneiwitten hebben alfa helices (ongeveer 20 AZ)
- sommige integrale membraaneiwitten hebben meer membraanspanning domeinen
- sommige integrale eiwitten zijn gelinkt met membraanfosfolipiden via een oligosaccharide of zijn
rechtstreeks gekoppeld aan een VZ of prenylgroep
Transmembraantransport: cytoplasme: ER -> proteïnen voor secretie en transmembraanproteïnen
• SRP (signal recognation particle) bindt op keten, ook GTP molecule bindt
• mRNA -> proteïne synthese start -> ribosoom dissocieert
• ribosoom gaat naar RER en geeft RNA af
• proteïne wordt aangemaakt en tegelijkertijd ook getransloceert
Localisatie signaal = signaal in mRNA of in gesynthetiseerde proteïne dat zegt aan ribosoom wat
soort proteïne wordt geproduceert en waar in de cel het thuishoort
4
, 8.2. Incorporeren van transmembraanproteïnen
AZ zijn + en – geladen
Grootste deel van transmembraanproteïnen zijn opgebouwd uit transmembraanhelices (alfa-helices)
die overspannen zijn door ladingen
1. Proteïnen zijn aangemaakt -> start transfer signaal
2. Eerste residu is negatief geladen AZ en zit in cytosol (ook – geladen)
3. Negatieve lading wordt afgestoten en naar de andere kan gebracht
4. Helix wordt gemaakt en wordt meestal door + AZ afgesloten
OF
1. Proteïnen zijn aangemaakt -> start transfer signaal
2. Eerste residu is een + geladen AZ
3. N-terminus blijft in de cytosol en C-terminus is volledig getransloceerd naar de extracellulaire
zijde
Apoptose = cellen verliezen assymetrie in plasmamembraan door een tekort aan ATP,
fosfatidylserine (EC) is een receptor voor de fagocytose
9. Membraanproteïnen zijn mobiel
1) Diverse functies (transport, signalling, cytoskelet koppeling)
2) Membraan associatie
a. Integrale membraanproteïnen bevatten transmembraan peptide segmenten
b. Lipide anker zoals myristylatie, prenylatie, glycofosfolipiden
c. Indirect: perifere membraanproteïnen zijn gebonden aan integrale
membraanproteïnen
Fluorescent membraan -> deel weglaseren ->
na verloop van tijd herstel
Besluit: bewijs voor mobiliteit van
membraanproteïnen
several spectroscopic techniques: single molecules
5
structuur
1. Oppervlak van de cel wordt gedefinieerd door een membraan
Opbouw:
• Fosfolipidendubbellaag door amfipatisch karakter:
o Hoofd = polair/ hydrofield (wijst naar het water)
o VZ staart = apolair/hydrofoob (wijst weg van het water)
• Ondoorlaatbaar voor grote moleculen (proteïnen en NZ)
• Selectief permeabel voor kleine moleculen (ionen en metabolieten)
• Actief transport doorheen het membraan = stoffen worden tegen hun concentratiegradiënt
in via een membraan-eiwit (ATP) opgenomen in de cel of afgestaan aan het milieu. (stoffen
worden dus opgenomen vanuit een gebied met een lage conc -> hoge conc)
2. Celmembraan is opgebouwd uit fosfolipiden
Fosfolipidenmembraan vormt in laag geconcentreerd waterig milieu een monolaag
En in een hoog geconcentreerd waterig milieu micellen
Bij een nog hogere concentratie aan water wordt een fosfolipidedubbelllaag gevormd = bilayer
Leaflets = fosfolipidemoleculen liggen in 2 parallelle rijen staart à staart tegen elkaar
Bilayer:
-> spontane vesikelvorming
-> spontaan helen bij ruptuur (inscheuren van weefsel)
-> self-sealing: kan geen vrije rand hebben
-> spontane opvouwing/regeneratie
1
,Meeste membraanlipiden = fosfolipiden (bv. fosfatiylcholine)
• Hoofd = hydrofiel = glycerol – fosfaatgroep – choline
• Staart = hydrofoob = 2 acylstaarten
• Verzadigd/ saturated= recht
• Onverzadigd/ unsaturated = knik t.h.v. dubbele binding
• Verzadigheid/onverzadigdheid beïnvloednen de VL, dikte en structuur van het membraan
• Fosfolipiden van dierlijke cellen bevatten meestal 1 verzadigde en 1 onverzadigde VZ staart
Andere membraanlipiden
- cholesterol
- glycolipiden: fosfatidylserine, fosfatidylinositol
- sfingolipiden (afkomstig van sfingosine) : glycosfingolipiden, sfingomyeline, gangliosides
3. Bilayer vormt een 2D vloeibare flase
Beweging van lipiden in de fosfolipidendubbellaag:
• Laterale diffusie = snel
• Rotatie/flexie van lipide = snel
• Flip-flop beweging van de ene -> andere leaflet = traag en gebeurd nauwelijks -> hoge
energie barrière verreist en floppase enzymen
Flipase = fosfolipiden van EC -> IC brengen
Flopase = fosfolipiden van IC -> EC brengen
Scramblase = fosfolipiden van EC -> IC en IC -> EC brengen (kan beide)
4. Vloeibaarheid van het membraan wordt bepaald door lipiden compositie
1. Zuiver fosfolipide membraan: transitietemperatuur hangt af van samenstelling
(packing) membraan:
• Lengte fosfolipiden staarten: 14-24C (meeste 18-20), hoe korter de staart -> hoe
vloeibaarder het zuiver fosfolipide membraan
• Aantal onverzadigdheden in lipide staarten:
o Verzadigd: recht -> dichte/sterke packing
o Onverzadigd: knik -> minder/zwakke packing -> vloeibaar bij lagere temperatuur
o Gevolg (nadeel) van zwakke packing is dat het membraan scheuren/gaten vertoont
en dus meer permeabel is
• Korte verzadigde ketens -> lage Tm (bv. olie)
• Lange onverzadigde ketens -> hoge Tm (bv. boter)
Tm = transitietemperatuur = temperatuur waarbij de
bilayer overgaat van de vaste ‘gel’ fase naar de vloeibare
‘sol’ (en omgekeerd)
2
, 2. Biologisch membraan:
• Zelfde algemene principes maar meer soorten lipiden
• Cholesterol intercaleerd met onverzadigde lipiden en heeft 2 effecten:
o Rigide ring stabiliseert C-atomen nabij ‘head’ -> rigider, minder permeabel
o Cholesterol verhindert staart-staart interactie (voorkomt kristallisatie van fosfolipide
staarten) -> membraan blijft vloeibaar
o Invloed op dikte van het membraan -> wordt dikker
hoge concentratie aan cholesterol -> vloeibaarheid membraan stijgt
in tegenstelling tot fosfolipiden gebeuren cholesterol
flipflops makkelijk over het membraan omdat
cholesterol een polair hoofdje heeft
-> gelijke concentratie aan cholesterol aan de binnen
en buitenkant van het membraan
Stijfheid/ vloeibaarheid van de bilayer bepaald de
permeabiliteit van het membraan
5. Fosfolipidedubbellaag is ondoorlaatbaar voor geladen moleculen
Zuivere fosfolipidedubbellaag is ondoorlaatbaar voor
-> ionen: Na+, K+, Cl-, Ca+
-> grote wateroplosbare moleculen: proteïnen, NZ, suikers en nucleotiden
Permeabel voor:
-> O2, CO2 en NH3
6. Asymmetrie in biologische membranen
Inner leaflet (intracellulair): Outer leaflet (extracellulair):
- fosfatidylinositol - glycolipiden
- fosfatidylethanolamine - sfingomyeline
- fosfatidylserine - fosfatidylcholine
Fosfatidylinositol -> betrokken bij second messenger signaal: IP3-DAG/PKC signaal cascade -> IC
Glycolipiden enkel aan de extracellulaire zijde: door synthese in ER/ golgi lumen
Fosfatidylchoine wordt in de binnenzijde aangemaakt en via flopase naar buitenzijde
3
,Aanmaak van fosfolipiden in SER of GA in lumen of in de cytoplasmatische zijde
Asymmetrie in fosfolipide compositie tussen inner en outer leaflet van het membraan:
• Ontstaan tijdens biosynthese (ER/golgi: flippase/floppase)
• Beïnvloed de buiging en vloeibaarheid van het membraan (EC is stijver dan IC)
• Cytosol leaflet (IC) is negatiever dan EC, reguleert incorporatie van membraan proteïnen
o Fosfatidylserine heeft een negatieve lading, de andere FL zijn neutraal
Specifieke compositie is soms nodig voor de functie van membraan proteïnen
7. Lipid rafts
Lipid raft = locale concentratie van specifieke lipiden, sfingomyeline, cholesterol en proteïnen
• Meestal dikker dan de rest van het membraan door aanrijking van verzadigde fosfolipiden
• Clusters van proteïnen/lipiden (rafts) spelen een rol in signaaltransductie (concentratie
second messengers zoals PIP3)
• Bevatten glycolipids (geglycosyleerde fosfolypiden
8. Correct incorporeren van transmembraanproteïnen
8.1. Transmembraanproteïnen
Soorten:
- perifere proteïnen zijn niet covalent gebonden met integrale proteïnen
- meeste integrale membraaneiwitten hebben alfa helices (ongeveer 20 AZ)
- sommige integrale membraaneiwitten hebben meer membraanspanning domeinen
- sommige integrale eiwitten zijn gelinkt met membraanfosfolipiden via een oligosaccharide of zijn
rechtstreeks gekoppeld aan een VZ of prenylgroep
Transmembraantransport: cytoplasme: ER -> proteïnen voor secretie en transmembraanproteïnen
• SRP (signal recognation particle) bindt op keten, ook GTP molecule bindt
• mRNA -> proteïne synthese start -> ribosoom dissocieert
• ribosoom gaat naar RER en geeft RNA af
• proteïne wordt aangemaakt en tegelijkertijd ook getransloceert
Localisatie signaal = signaal in mRNA of in gesynthetiseerde proteïne dat zegt aan ribosoom wat
soort proteïne wordt geproduceert en waar in de cel het thuishoort
4
, 8.2. Incorporeren van transmembraanproteïnen
AZ zijn + en – geladen
Grootste deel van transmembraanproteïnen zijn opgebouwd uit transmembraanhelices (alfa-helices)
die overspannen zijn door ladingen
1. Proteïnen zijn aangemaakt -> start transfer signaal
2. Eerste residu is negatief geladen AZ en zit in cytosol (ook – geladen)
3. Negatieve lading wordt afgestoten en naar de andere kan gebracht
4. Helix wordt gemaakt en wordt meestal door + AZ afgesloten
OF
1. Proteïnen zijn aangemaakt -> start transfer signaal
2. Eerste residu is een + geladen AZ
3. N-terminus blijft in de cytosol en C-terminus is volledig getransloceerd naar de extracellulaire
zijde
Apoptose = cellen verliezen assymetrie in plasmamembraan door een tekort aan ATP,
fosfatidylserine (EC) is een receptor voor de fagocytose
9. Membraanproteïnen zijn mobiel
1) Diverse functies (transport, signalling, cytoskelet koppeling)
2) Membraan associatie
a. Integrale membraanproteïnen bevatten transmembraan peptide segmenten
b. Lipide anker zoals myristylatie, prenylatie, glycofosfolipiden
c. Indirect: perifere membraanproteïnen zijn gebonden aan integrale
membraanproteïnen
Fluorescent membraan -> deel weglaseren ->
na verloop van tijd herstel
Besluit: bewijs voor mobiliteit van
membraanproteïnen
several spectroscopic techniques: single molecules
5