Cellen en weefsels samenvatting H.13
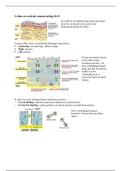
In de cellen is er verschillend vesiculair transport. Je hebt secretie (rood), endocytose (groen)
en terugkerende paden (blauw). De vesikels bewegen zich voort door motoreiwitten. Als een
vesikel van organel verplaatst dan zijn er drie fases: diffusie, budding en fusie. Door het
vesiculair transport verliezen de organellen hun identiteit niet (golgi blijft golgi).
Om het membraan van een vesikel zit een coat. Deze coat heeft als functie om cargo
moleculen te selecteren en om de membraan buiging te stabiliseren. De drie meest
voorkomende coats zijn: clathrin, COPI en COPII. Alle pijltjes die in het plaatje te zien zijn,
worden gemedieerd door microtubuli.
De coat tussen het golgi en het plasmamembraan is de clathrin coat. Deze coat bestaat uit 6
en 5 hoeken (net zoals een voetbal). De coat is opgebouwd uit triskelions. Als er een ligand is
gebonden, dan verandert de receptor van conformatie en gaat deze binden aan adaptor
eiwitten. Hierdoor gaat het membraan buigen. De clathrin kan binden aan de adaptoreiwitten
en zorgt voor stabilisatie, en dit is na de vorming niet meer nodig (daarom dissociëren ze). De
adaptor eiwitten selecteren dus voor cargo moleculen en de buigen van het membraan wordt
mogelijk gemaakt door clathrin triskelions en BAR domeinen.
In het membraan zitten speciale lipiden, inositol lipiden. In verschillende
delen van de cel zitten verschillende fosfatidylinositol lipiden waardoor de
organellen en membraan domeinen ‘gemerkt’ worden. Aan deze lipiden
binden adaptormoleculen (herkennen door speciale lading kopgroep).
, Het AP2 adaptor eiwit bestaat uit vier domeinen. Als het bindt aan de PI(4,5)P2 lipiden in het
membraan, en verandert de conformatie zodat de binding sites voor de receptoren vrij komen
te liggen. Deze binding sites binden aan receptoren als deze de juiste endocytotische signalen
uitzenden (als niet zo is, dan dissociëren AP2 complexen weer). Dit is een sterke binding
waardoor het membraan buigt. Dit is voordelig voor nabijgelegen AP2 complexen om ook te
binden. De buiging voor het membraan wordt gestabiliseerd door BAR domeinen (een soort
van banaan eiwitten). Deze hebben een hydrofoob staartje wat in het membraan zit, waardoor
het vast blijft zitten. Elke keer als de vesikel meer vormt, buigt het membraan meer.
Als de vesikels klaar zijn, moeten ze worden afgesnoerd van het membraan. Dit gebeurt
m.b.v. het eiwit dynamine. Dynamine is een GTPase en GTP wordt gehydrolyseert, dan
verandert het eiwit van conformatie. Door deze conformatie verandering wordt het membraan
als het ware gewurgd, en komen de membranen zo dicht bij elkaar dat ze fuseren.
Als de vesikels los zijn, dan moet de coat verwijdert worden. Eerst
worden de PI94,5)P2 lipiden gehydrolyseert door een fosfatase
waardoor de AP2 binding verzwakt. Daarna wordt de coating eraf
gehaald door ATPases (hsp70 chaperonnes).
Er kunnen naast clathrin coated vesikels ook COPII
coated vesikels gemaakt worden. Deze worden vaak
gemaakt in het ER. Hierbij bindt inactief Sar1-GDP aan
Sar1-GEF in het membraan, waardoor GDP wordt
vervangen. Hierdoor vindt er een conformatie
verandering plaats waardoor er een alfa helix in het
membraan gaat en Sar1-GTP dus in het membraan zit.
Sar1-GTP bindt aan Sec23 en Sec24 als een cargo
molecuul is gebonden en samen vormen ze de
binnenkant van de coat (het complex zorgt ook voor de
membraan buiging). Actief Sar1-GTP zorgt ook voor de
aantrekking van COPII coat eiwitten Sec13/31.
In de cellen is er verschillend vesiculair transport. Je hebt secretie (rood), endocytose (groen)
en terugkerende paden (blauw). De vesikels bewegen zich voort door motoreiwitten. Als een
vesikel van organel verplaatst dan zijn er drie fases: diffusie, budding en fusie. Door het
vesiculair transport verliezen de organellen hun identiteit niet (golgi blijft golgi).
Om het membraan van een vesikel zit een coat. Deze coat heeft als functie om cargo
moleculen te selecteren en om de membraan buiging te stabiliseren. De drie meest
voorkomende coats zijn: clathrin, COPI en COPII. Alle pijltjes die in het plaatje te zien zijn,
worden gemedieerd door microtubuli.
De coat tussen het golgi en het plasmamembraan is de clathrin coat. Deze coat bestaat uit 6
en 5 hoeken (net zoals een voetbal). De coat is opgebouwd uit triskelions. Als er een ligand is
gebonden, dan verandert de receptor van conformatie en gaat deze binden aan adaptor
eiwitten. Hierdoor gaat het membraan buigen. De clathrin kan binden aan de adaptoreiwitten
en zorgt voor stabilisatie, en dit is na de vorming niet meer nodig (daarom dissociëren ze). De
adaptor eiwitten selecteren dus voor cargo moleculen en de buigen van het membraan wordt
mogelijk gemaakt door clathrin triskelions en BAR domeinen.
In het membraan zitten speciale lipiden, inositol lipiden. In verschillende
delen van de cel zitten verschillende fosfatidylinositol lipiden waardoor de
organellen en membraan domeinen ‘gemerkt’ worden. Aan deze lipiden
binden adaptormoleculen (herkennen door speciale lading kopgroep).
, Het AP2 adaptor eiwit bestaat uit vier domeinen. Als het bindt aan de PI(4,5)P2 lipiden in het
membraan, en verandert de conformatie zodat de binding sites voor de receptoren vrij komen
te liggen. Deze binding sites binden aan receptoren als deze de juiste endocytotische signalen
uitzenden (als niet zo is, dan dissociëren AP2 complexen weer). Dit is een sterke binding
waardoor het membraan buigt. Dit is voordelig voor nabijgelegen AP2 complexen om ook te
binden. De buiging voor het membraan wordt gestabiliseerd door BAR domeinen (een soort
van banaan eiwitten). Deze hebben een hydrofoob staartje wat in het membraan zit, waardoor
het vast blijft zitten. Elke keer als de vesikel meer vormt, buigt het membraan meer.
Als de vesikels klaar zijn, moeten ze worden afgesnoerd van het membraan. Dit gebeurt
m.b.v. het eiwit dynamine. Dynamine is een GTPase en GTP wordt gehydrolyseert, dan
verandert het eiwit van conformatie. Door deze conformatie verandering wordt het membraan
als het ware gewurgd, en komen de membranen zo dicht bij elkaar dat ze fuseren.
Als de vesikels los zijn, dan moet de coat verwijdert worden. Eerst
worden de PI94,5)P2 lipiden gehydrolyseert door een fosfatase
waardoor de AP2 binding verzwakt. Daarna wordt de coating eraf
gehaald door ATPases (hsp70 chaperonnes).
Er kunnen naast clathrin coated vesikels ook COPII
coated vesikels gemaakt worden. Deze worden vaak
gemaakt in het ER. Hierbij bindt inactief Sar1-GDP aan
Sar1-GEF in het membraan, waardoor GDP wordt
vervangen. Hierdoor vindt er een conformatie
verandering plaats waardoor er een alfa helix in het
membraan gaat en Sar1-GTP dus in het membraan zit.
Sar1-GTP bindt aan Sec23 en Sec24 als een cargo
molecuul is gebonden en samen vormen ze de
binnenkant van de coat (het complex zorgt ook voor de
membraan buiging). Actief Sar1-GTP zorgt ook voor de
aantrekking van COPII coat eiwitten Sec13/31.